Жировая ткань – I
Жировая ткань является одной из самых распространенных тканей в организме и составляет около 15-20% от веса тела у мужчин и 20-25% у женщин. 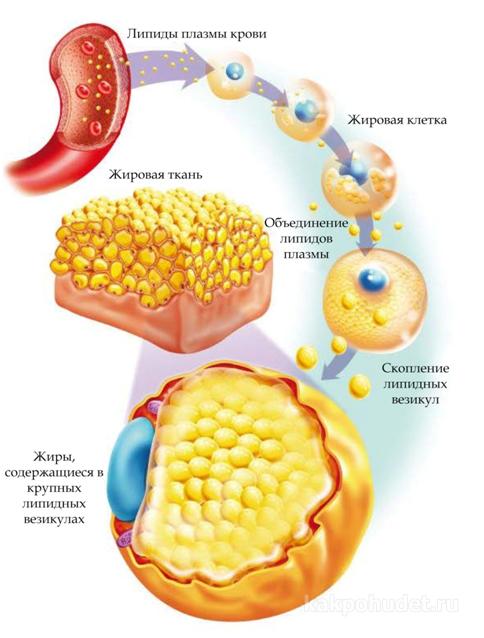 Это особый вид соединительной ткани с преобладанием клеток, называемых адипоцитами, которые очень эффективно хранят энергию в виде триглицеридов.
Это особый вид соединительной ткани с преобладанием клеток, называемых адипоцитами, которые очень эффективно хранят энергию в виде триглицеридов.
Жировая ткань играет незаменимую роль в поддержании энергетического баланса в организме млекопитающих. В периоды перенасыщения, она способна хранить энергию в форме триглицеридов, а затем выпускать эту энергию в виде свободных жирных кислот в условиях недостатка калорий.
Это не просто «хранилище энергии»; было обнаружено, что она ведет себя как эндокринный орган, синтезируя и выделяя молекулы, связанные с регулированием потребления и расхода энергии (лептин, АсгрЗО/AdipoQ), сосудистым гомеостазом (ангиотензин, ингибитор активатора плазминогена 1 типа), и иммунным ответом (TNF-альфа).
Поэтому не удивительно, что изменения в физиологии и метаболизме жировых тканей могут быть связаны с такими нарушениями как ожирение, дислипидемия, инсулинорезистентность и сахарный диабет 2 типа. В зависимости от характеристик клеток, составляющих ее основу, жировую ткань можно разделить на однокамерную или многокамерную.
Жировая ткань – II
Однокамерный тип, также именуемый «белым жиром» – это жировая ткань, которая, как правило, встречается у взрослых; она обладает сильной васкуляризацией и представляет совокупность полиэдральных клеток (адипоцитов), которые составляют 10-100 микрон в диаметре (и которые могут достигать до 200 микрон при некоторых формах ожирения).
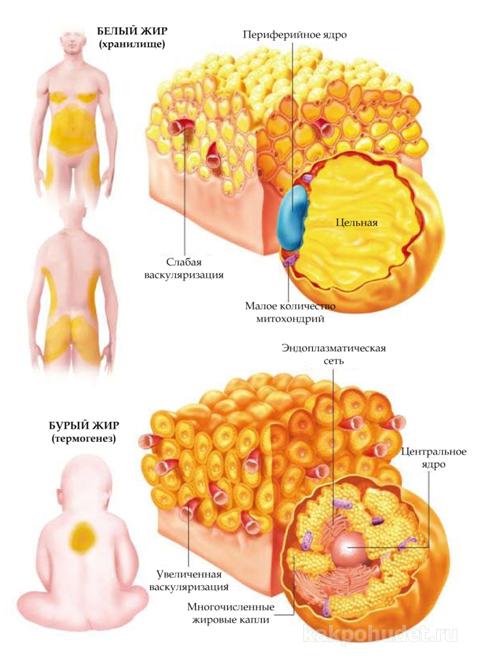
Адипоциты содержат крупные капли жира, которые занимают все цитоплазматическое пространство и вытесняют органеллы и ядро к периферии. При световой микроскопии можно наблюдать «перстеневидную» форму клеток, а при электронной микроскопии просматривается цельная липидная капля с перинуклеарной цитоплазмой, в которой содержится небольшой аппарат Гольджи, микротрубочки и пиноцитозные пузырьки, редкие митохондрии овальной формы, относительно слабо развитый шероховатый эндоплазматический ретикулум и свободные рибосомы.
Многокамерная жировая ткань встречается во взрослом организме в ограниченном количестве, хотя она в изобилии присутствует у зародышей и новорожденных детей (в межлопаточной области). Ее клетки имеют полигональную форму и сравнительно меньшие размеры, нежели клетки однокамерной жировой ткани, а в цитоплазме можно наблюдать многочисленные жировые капли различных размеров.
Ядро, как правило, расположено в центре и имеет сферическую форму, присутствуют многочисленные митохондрии, с хорошо развитыми складками-кристами. Этот тип жировой ткани, еще более васкуляризированный, чем однокамерный, также известен как «бурый жир».
Происхождение адипоцитов
Эмбриональное происхождение жировых клеток до конца не изучено, хотя многие исследования показывают, что линия адипоцитов происходит 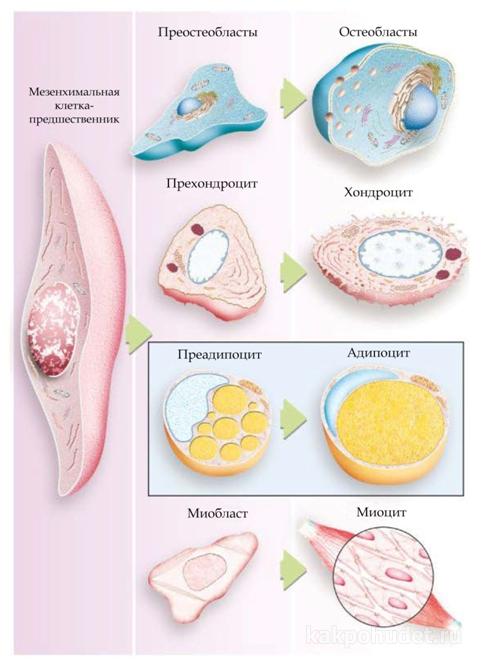 из тотипотентных стволовых клеток, которые могут дифференцироваться в унипотенциальные клетки, которые, в свою очередь, развиваются (путем еще плохо изученных молекулярных механизмов) в специфические типы клеток, такие как хондробласты, миобласты и адипобласты.
из тотипотентных стволовых клеток, которые могут дифференцироваться в унипотенциальные клетки, которые, в свою очередь, развиваются (путем еще плохо изученных молекулярных механизмов) в специфические типы клеток, такие как хондробласты, миобласты и адипобласты.
Преадипоциты (морфологически схожие с фибробластами) – это клетки, призванные дифференцироваться в клеточную линию адипоцитов. Они развиваются в клетки сферической формы, которые начинают накапливать липиды и это запускает процесс развития морфологических и биохимических характеристик зрелых адипоцитов. Этот процесс дифференциации является в высшей степени сложным и управляется различными гормонами и факторов роста.
После дифференциации «зрелые» адипоциты теряют способность к делению; однако это клетки с очень большим периодом полураспада и они могут существенно увеличивать свой липидный запас. Свежесформированная жировая ткань содержит остаточные неразвившиеся адипоциты и стволовые клетки, которые могут дифференцироваться в дополнительные адипоциты в случае чрезмерного потребления калорий.
Метаболизм адипоцитов – I
В настоящее время под адипоцитом понимается клетка, активно регулирующая различные метаболические пути, ответственные за энергетический баланс, и чья деятельность контролируется сложной системой нейрогормональных сигналов. 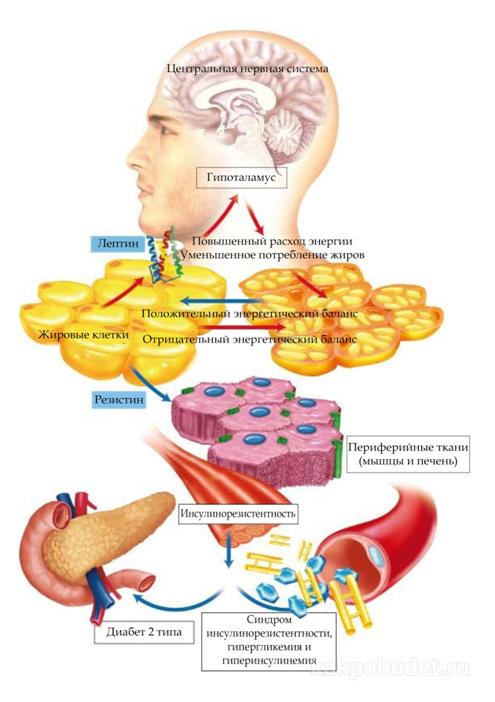 Существует двунаправленная связь между адипоцитами и другими тканями, ввиду того, что большое количество веществ, выделяемых жировой тканью (в общих смысле именуемых адипокинами) встроено в химическую систему эккринных и паракринных сигнальных путей, многие из которых до сих пор полностью не изучены.
Существует двунаправленная связь между адипоцитами и другими тканями, ввиду того, что большое количество веществ, выделяемых жировой тканью (в общих смысле именуемых адипокинами) встроено в химическую систему эккринных и паракринных сигнальных путей, многие из которых до сих пор полностью не изучены.
Некоторые из секретируемых пептидных продуктов, таких как TNF-альфа и резистин, могут вызвать состояние периферической резистентности к инсулину, тогда как другие, например, лептин и адипонектин, способствуют развитию восприимчивости к инсулину. Лептин, выделяемый наполненными триглицеридом адипоцитами, достигает гипоталамуса через общий кровоток, пересекая гематоэнцефалический барьер, снижая потребление калорий и увеличение их расход посредством модуляции нейроэндокринной и вегетативной систем.
Адипонектин (AdipoQ или АсргЗО) – это адипокин, синтезируемый и выделяемый исключительно адипоцитами; недавно было выявлено, что он влияет на гомеостаз глюкозы и восприимчивость к инсулину. Кроме того, было также обнаружено, что адипоциты могут секретировать белки комплементарной системы, провоспалительные, прокоагулянтные цитокиноы и стресс-белки.
Метаболизм адипоцитов – II
Циркулирующие в крови хиломикроны и ЛПОНП являются основным источником триглицеридов (ТГ) — жиров, хранящихся в адипоцитах. Липопротеинлипазы (ЛПЛ) стенок капилляров жировой ткани гидролизуют ТГ липопротеинов до свободных жирных кислот (СЖК) и глицерина. 
Адипоциты впитывают СЖК посредством активных транспортных процессов при участии специфических транспортных белков жирных кислот. Внутри клетки СЖК реэтерифицируются с образованием ТГ, которые накапливаются в качестве резервных источников энергии в период достаточного поступления калорий.
ЖК, циркулирующие в привязке к альбумину или синтезирующиеся в клетке из глюкозы «с нуля», проходят аналогичный процесс в ситуации избыточного потребления углеводов.
В процессе липолиза ТГ, хранящиеся в адипоцитах, гидролизуются до жирных кислот и глицерина. Ингибирующим ферментом этого метаболического пути является гормон-чувствительная липаза (ГЧЛ), которая катализирует разложение триглицеридов до моноглицеридов, гидролизующихся моноацилглицероль-липазой.
ГЧЛ – это гормон со строгой регуляцией; он активируется путем фосфорилирования посредством циклической АМФ-зависимой протеинкиназы. Липолиз стимулируется всеми теми гормонами, которые в привязке к своим рецепторам приводят к стимулированию активации G белков, образования циклического АМФ и активации ГЧЛ (катехоламины, глюкагон, соматотропин, глюкокортикоиды). Гормоны, ингибирующие липолиз (такие как инсулин), активируют рецепторы, связанные с фосфатидилинозит 3-киназой и таким образом активируют фосфодиэстеразу 3, которая катализирует превращение цАМФ в 5’АМФ.